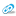
Всё гениальное беспредельно по смыслу и нужно это только заметить и отследить. Нас греет печень, и даже филология здесь подсказка (печка, т.е. печень). В ней «несметное» (никто точно не определил до сих пор) число гепатоцитов, в каждом 1560 митохондрий и 20 тыс. дыхательных ансамблей (дыхательных цепей, цепей переноса электронов во всей структурно-молекулярной сложности). А по сути биологической (физиологической, метаболической, химической, физической, волновой – всё это обозначение молекулярно-организменной вертикали) - это «место встречи (которое изменить нельзя») кислорода (извне) и водорода (изнутри) для управляемого синтеза воды. В сутки мы синтезируем в покое 0,5 л воды. При лихорадке образуется больше и бесконечно (кому как повезет в этой жизни в состоянии болезни!). Синтез воды (т.е. реакция «гремучего газа» по Лавуазье 2Н2 + О2 = 2Н2О + 56 ккал) образует энергию в виде тепла и для огромнейшего числа метаболических реакций в виде «разменной валюты», т.е. АТФ (АДФ+Ф+10ккал=АТФ), когда дозировано нужно получить и доставить энергию для конкретной реакции синтеза или обмена. Разумеется, для этого нужно много фосфата, т.е. Н3РО4. Им нужно не только орошать, но и пропитывать все закоулки организма и самыми дальними и многочисленными являются митохондрии. Дышим мы (не ртом как индеец в «Следопыте» у Ф.Купера и не грудью как дама из «Нетерпения сердца» у С.Цвейга) митохондриями и это прямая вселенско-организменная связь. Поэтому фосфаты лучше иметь не только в достатке, но и избытке. Тут-то и появляется ЩФ со своей широчайшей специфичностью, а точнее неспецифичностью и она «отгрызает фосфаты от любого метаболита, чтобы пропитать им весь организм. Иначе «дежурит» на входе в биоэнергетику, когда рассеивающееся тепло приобретает конкретно-жизненный смысл, как и печь, на которой нужно ещё и что-то приготовить. Не случайно эта величина константна для каждого возраста: в раннем детстве – 400 МЕ/л (когда траты еще безумны и не отрегулированы), а к моменту созревания (хотя бы полового) – 100 МЕ/л. Поэтому гениальные строки А.Суркова полностью отражают суть всего живого: огонь «бьется» и в митохондриях, но необходимо не только тепло, но и его консервация для многочисленных житейских (то бишь метаболических) нужд. А без фосфата это невозможно. Поэтому «смола, как слеза» это не только поэтический, но и равновеликий образ всего сущего. Можно продолжать и дальше, но …… объем издания тормозит и даже душит как удавка.
Важность ЩФ в характеристике метаболизма связана с ее центральной ролью в поддержании глюкозо-фосфатного гомеостаза. ЩФ – осуществляет процессы трансмембранного фосфорилирования, обеспечивая вход и выход глюкозы в клетки, что напрямую влияет на уровень глюкозы в крови, соответственно также играет роль в поддержании уровня фосфатов, необходимых для биоэнергетики.
Интегративная схема метаболизма в физиологической интерпретации с комментариями:
Главная биоэнергетическая реакция: 2Н2 + О2= 2Н2О56 ккал.
Сгорание глюкозы в организме: С6Н12О6+6О2 = 6СО2 (в общих путях катаболизма) + 6Н2О (управляемый синтез в общих путях катаболизма) 686 ккал.
Глюкоза + АТФ = Глюкозо-6-фосфат: стартовая реакция в обмене углеводов, начало кинеза (отсюда и название фермента гексокиназа) в метаболизме, запирающая в клетке в фосфорилированном виде глюкозу реакция (заряды фосфорной кислоты мешают выходу глюкозы из нее).
Этапы горения глюкозы в организме в водной среде и при 36°6С: 1) гликолиз, 2) 1-й общий путь катаболизма (окислительное декарбоксилирование пирувата) – первый выход СО2, 3) 2-й общий путь катаболизма (цикл трикарбоновых кислот или цикл Кребса) – второй выход СО2 и образование водородов для синтеза воды, 4) 3-й общий путь катаболизма (цепь тканевого дыхания или цепь переноса электронов) – место управляемого синтеза воды.
Маркируются вышеописанными ферментами интенсивность: ЦТК – АСТ, ГНГ – АЛТ, гликолиза – ЛДГ, энергорасхода – КФК, трансмембранных переходов фосфата – ЩФ, транспорта аминокислот – ГГТ.

ЩФ – осуществляет процессы трансмембранного фосфорилирования, обеспечивая вход и выход глюкозы в клетки, что напрямую влияет на уровень глюкозы в крови, соответственно также играет роль в поддержании уровня фосфатов, необходимых для биоэнергетики.
Биоэнергетическая машина клетки имеет несколько уровней регуляции. Первый уровень субстратной регуляции обеспечивается исходными метаболитами для синтеза и транспорта макроэргов, к которым относится креатин. Креатин своим гидрофильным «концом», имеющим высокий энергетический потенциал извлекает из митохондрий заряженный фосфат от АТФ с сохранением в них АДФ для последующего ресинтеза. Именно поэтому его концентрация в крови низкая, стабильная и фактически регулирует биоэнергетику организма. Синтез креатина имеет следующие особенности: во-первых, необходим «вездесущий» глицин, во-вторых, «дефицитные» аргинин и метионин. На первом этапе гианидиновая группа аргинина переносится на глицин при участии фермента трансамидиназы (Е.С.2.6.2.1), которая находится в конкурентных взаимоотношениях с аргиназой из цикла синтеза мочевины, т.к. синтез креатина и мочевины метаболически взаимосвязан. Много мочевины – мало креатина. Дефицит аргинина тормозит синтез креатина. Поэтому в клинике нередко отмечается мочевино-креатининовая диссоциация как показатель жесткой регуляторной зависимости. Этот механизм компенсирует избыток мочевины. Переход креатина в креатинин можно рассматривать и как консервацию энергии в митохондриях с отключением механизма извлечения АТФ. Первая реакция синтеза обратима и при рН 8,5 протекает в обратном направлении. Характерно, что активность трансамидиназы зависит от пула глутатиона и при его окислении (переход сульфгидрильной группы в дисульфидную) активность фермента снижается. Креатинфосфокиназа (КФК) является индикатором интенсивности использования доступного креатина и ее активность определяется двумя факторами: во-первых, содержанием креатина, во-вторых, интенсивностью синтеза и разрушения макроэргов (АТФ и КФ).
Второй этап необратим (регуляция заканчивается и наступает работа)и протекает в поджелудочной железе и печени путем реакции метилирования гуанидинацетатметилтрансферазой (ГАМТ-2.1.1.2) при участии активированной формы метионина (S-аденозилметионина). В активном центре ведущее значение придают четырем тиоловым группам: Cys-15, Cys-90, Cys-207, Cys-219, которым необходимо «глутатионовое орошение» и их окисление, поэтому направленный мутагенез вызывают снижение активности фермента. Состояние активного центра зависит и от пула глутамина, который характеризует состояние белкового обмена (фактический акцептор избытка аммиака) – он восстанавливает GSSG инактивированный фермент [A6]. Присоединение метильной группы усиливает энергетический заряд гидрофильного «наконечника» креатина, что способствует «вытаскиванию» макроэргических фосфатов из митохондрий. Аргинин является не только предшественником креатина, но и является источником синтеза NO, который регулирует местную гемодинамику и от которой зависит активность КФК, т.е. аргинин является регулятором адекватного кровоснабжения соответственно уровню креатина. В данном случае наблюдается каталитическая специфичность – на один субстрат – аргинин – действуют три фермента (аргиназа, трансамидиназа) и более элементарно метаболический путь определяется: 1) константой Михаэлиса – степень сродства к субстрату, 2) компартментализацией и 3) регуляцией конечными продуктами.
глицином для начала бесчисленных синтезов – гема, креатина, глютатиона и т.д. – является метаболической первоосновой различных форм жизни. Образно говоря это как выход на сушу древних – был важнейшим эволюционным шагом. Также в филогенетическом отношении - это важнейший шаг в сторону полифункциональности и не случайно поэтому замена одного из атомов водорода в глицине ведет к созданию всего многообразия аминокислот, т.е. физиологической полифункциональности. Но самое главное заключается в том, что глицину отводится важнейшая роль в структурном обеспечении всей биоэнергетики. В тоже время в метаболическом отношении глицин не может быть в дефиците, т.к. его источником служит серин, треонин, из которого также образуется гормоноподобный ацетальдегид и главным источником является 3-глицерофосфат - промежуточный метаболит обмена липидов и углеводов. Это ведь и превентивный стимул для ускоренного сжигания избытка белков в митохондриях особенно после тяжелых внутрисосудистых катастроф обычно воспалительного генеза. Описано образование глицина из аммиака, углекислого газа и НАДН+Н+ при участии тетрагидрофолиевой кислоты в митохондриях при участии глицинсинтазы. Это не что иное как сохранение продуктов катаболизма при старте в анаболизм. При этом анаболические пути в обмене глицина превалируют над катаболическими – распад возможен только через серин. Глицин является хорошим буферным растворителем, глутатион стабилизатором белково-ферментных систем. Субстрат-субстратная подчиненность.
Второй уровень регуляции биоэнергетики связан с полифункциональными эффектами креатина. «Консервация» креатина путем перевода его в креатинин тормозит синтез АТФ, но усиливает теплопродукцию за счет свободного окисления при сжигании избытка аминокислот. Высокая активность КФК в этот период повышает креатинфосфат для восстановления лабилизированных мембран – продукция эндогенного неотона. При разрушении креатина с помощью креатиназы образуется мочевина и саркозин (метилглицин).Важнейшими эффектами следует считать действие креатинина на регуляцию собственного синтеза и работу цикла синтеза мочевины (формирование синдрома мочевино-креатининовой диссоциации).
Важность ЩФ в характеристике метаболизма связана с ее центральной ролью в поддержании глюкозо-фосфорного гомеостаза. Одновременно необходимо помнить, что это один из способов удаления фосфатов в энергетически спокойном состоянии и поэтому минимальные значения КФК отмечаются при максимальном значении креатинина, который дефицитен не только по происхождению (для синтеза необходим аргинин и метионин), но и в физиологическом отношении, т.к. именно он обеспечивает выход макроэргов из митохондрий, а значит способен тормозить лихорадочную реакцию. На фоне липидно-фосфатной диссоциации (БЛП снижаются, а ЩФ повышается) параллельно (табл.10) меняется активность ГГТ и коэф. де Ритиса при стабильности остальных показателей. ЩФ и ГГТ являются мембранными ферментами, а метаболизм БЛП зависит от состояния их рецепторного аппарата. Это происходит на фоне роста коэф. де Ритиса, что является абсолютно благоприятным признаком с точки зрения метаболизма – происходит выравнивание метаболических параметров. Умеренная интоксикация требует и незначительных энзимологических сдвигов.

Динамика изменения активности ЩФ и ГГТ абсолютно идентичная и это может быть связано с усилением трансмембранных процессов, тем более что фосфорилирование нативных белков является важнейшим регуляторным механизмом основных внутриклеточных процессов.
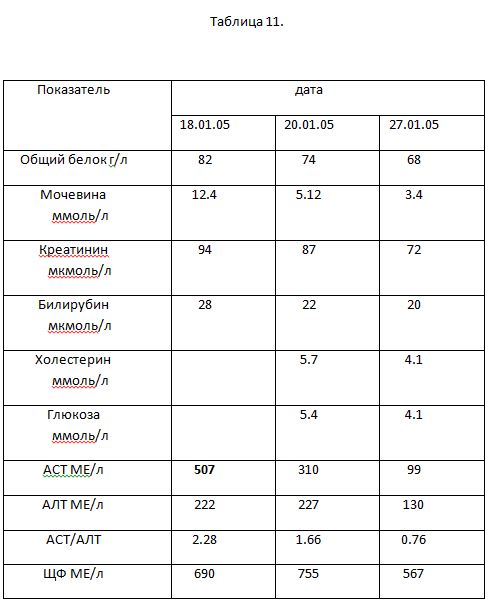
Все описанные выше состояния можно рассматривать как адаптивные реакции метаболического характера, которые метаболически жестко коррелируют и не отвечают требованиям однообразно направленного цитолиза. Абсолютно иной характер биохимических изменений наблюдается у больных ПТИ с делириозным статусом. В качестве примера приводим динамику больного (табл.11) С., история болезни № 311, 30 лет, поступившего во 2 КИБ г. Москвы. Заболел 14.01.05 после злоупотреблением алкоголем (пил водку).