





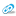
КФК ПРОДУЦЕНТ МЕМБРАНОПРОТЕКТОРА КРЕАТИНФОСФАТА (НЕОТОНА)
ИНДИКАТОР ЭФФЕКТИВНОСТИ ЭНЕРГЕТИЧЕСКИХ СДВИГОВ
СТИМУЛЯТОР АГРЕГАЦИИ ТРОМБОЦИТОВ
КОМПОНЕНТ ФОСФАТНОЙ БУФЕРНОЙ СИСТЕМЫ
ИНДИКАТОР ФОСФОЛИПАЗНОГО МЕХАНИЗМА ЦИТОЛИЗА И Т. Д.
Каждый слой представленной физиологической «иконы» фермента требует отдельного рассмотрения в цикле лекций и клинических примерах, но….нам всегда некогда и мы отвлекаемся на частности. «Феномен разобщения» активности АСТ и КФК нами описан 30 лет назад и суть его заключается в том, что при высокой КФК и низкой АСТ больные выживают (реанимация в инфекции), а при высокой АСТ и низкой КФК больные умирают. Задолго до летального исхода у больных вирусными гепатитами (повышение КФК для них нехарактерно) отмечается агональный выброс КФК. И это следствие катализируемой им в крови реакции: АТФ+креатин=АДФ+креатинфосфат (неотон по справочнику Видаля). КФК является важнейшим мембранопротектором, так как образует известное метаболическое средство – КФ или «неотон». Этот фермент влияет на фонд АДФ (индуктор агрегации тромбоцитов), а значит - на реологические свойства крови. В покое КФК может не определяться (ее активность является почти нулевой). Но при любом стрессе активность КФК повышается и в клинике практически до неограниченных (более 1000 МЕ/л) значений. КФК – абсолютно стресс-зависимый фермент, который является индикатором реализуемого энергетического потенциала организма.
Повышение в крови КФК, например, является информативным тестом шумовибрационного влияния на организм и это сопровождается дефицитом витаминов С, Р, В6, РР и возрастанием степени проницаемости эритроцитарных и других мембран, что отмечено у рабочих и подтверждено экспериментально на крысах.
КФК как компонент энергопродуцирующей системы различных тканей (мозг, сердце, мышцы): 1) катализирует как прямую, так и обратную реакции (синтез и распад креатинфосфата) с фармакологическими свойствами последнего, 2) реагирует на гормональные воздействия быстрого (адреналин) и длительного (глюкокортикоиды) характера, 3) чувствительна к детергентному для мембран и разобщающему для биологического окисления действию свободных жирных кислот, 4) относится к лабильной энергопродуцирующей системе по сравнению с инертной АСТ. Фактически вся система КФК является системой адаптации к изменяющимся биоэнергетическим потребностям миокарда, мозга и мышц при разном уровне кровоснабжения. Находится в конкурентных или синергичных взаимоотношениях с ЩФ за неорганический фосфат и нарабатывает АДФ – регулятор агрегации тромбоцитов. КФК-система работает в следующих ключевых зонах метаболизма: 1) регулирует соотношениея АТФ/АДФ в клетке, влияя тем самым на процессы биологического окисления и гликолиза, 2) обеспечивает транспорт макроэргического фосфата из митохондрий в цитоплазму, 3) энергетически обеспечивает транспортные АТФазы, 4) регенерирует АТФ для Са-АТФаз и Са-зависимых органелл и механизмов, 5) транспортирует иРНК из ядра, 6) обеспечивает внутриклеточный баланс фосфора. С учетом этих позиций более высокие уровни КФК у крыс, например, объясняются и более высокими физиологическими характеристиками: пульс и частота дыхания. Физиологические параметры жестко константны у человека в виде дыхательной эуритмии и выражаются следующей формулой: 20 (число дыханий) х 4 = 80 (пульс) х 4 = 320 (скорость диссоциации Нb), а у крыс с более коротким сроком жизни и иными физиологическими параметрами требуются и более вариативные адаптивно-пригодные метаболические системы. Крысы выживают популяционно, а человек индивидуально. И интеллектуальная надстройка требует высокого уровня белка (у крыс 65 г/л, а у человека 75 г/л при более низком термогенезе: у крыс АСТ 400 МЕ/л, а у человека 30 МЕ/л).
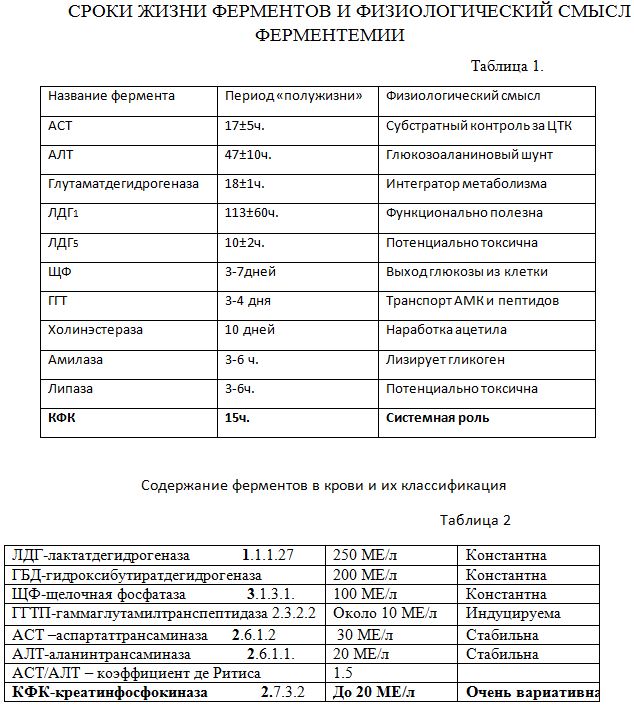
Обращает внимание разноуровненность ферментов, которая зависит от характера и интенсивности катализируемой реакции в метаболических путях. КФК как бы завершает оба ряда по минимальным срокам жизни и ферментативной активности, но ….. только в абсолютном покое (состояние близкое к нирванне). КФК-система это МЧС в организме и подключать её очень дорого и требуется уникальная оптимальность на всех организменных этажах в молекулярно-организменной вертикали и альвеолярно-митохондриальной горизонтали. Это тяжкий крест понимания сущности адаптации, который только и может помочь избежать другого креста! Это сфокусированная суть всего живого, как винчестер компьютера. Вышестоящие ферменты обеспечивают примитивную, фундаментальную физиологию, а КФК помимо этого в крови способна мощно и длительно прибавить на все случаи жизни и все патологические состояния. «Счастье по-моему просто бывает разного роста, от кочки и до Казбека в зависимости от человека!» (Э.Асадов). И здесь кому как повезет с мощным ферментом спасения и адаптации, который неожиданно включается в борьбу за жизнь. Неотон в клинике вводят в граммовых количествах, фактически струйно.
КФК-система, состоящая как из лабильного фермента (КФК), так и константного субстрата (креатин). Креатин синтезируется из незаменимых аминокислот: аргинина и метионина и ключевого структурного метаболита глицина. Креатин метаболически полифункционален и обладает свойствами гормоно-подобного метаболита-регулятора как на уровне клетки, так и на уровне целостного организма с восстановительными свойствами для поврежденных мембран. У крыс отмечается в среднем стократное превышение уровня активности КФК по сравнению с человеком, и имеет максимальный диапазон колебаний (28-10000 МЕ/л), что соответствует ее биологической роли в качестве компонента энергопродуцирующией КФК-системы. При малой массе тела, низкой деградации креатина и высоком уровне активности КФК энергообеспечение через креатинфосфат более постоянно. КФК-система у человека находится в латентном состоянии и включается лишь под действием гормональных факторов на короткое время. Низкий уровень креатинина у крыс можно объяснить тем, что КФК-система работает постоянно при высоком уровне КФК, что требует большей концентрации креатина, а не продукта его деградации. Работа КФК-системы позволяет переходить на аэробный гликолиз, минуя анаэробное окисление с низкой эффективностью при накоплении кислых продуктов. Высокий уровень ЛДГ может быть «функционально» полезным при более интенсивном газообмене и гликолизе в эритроцитах, число которых почти вдвое выше (7.2-9.6 млн/мл), чем у человека. Подтверждением этого можно считать и высокие уровни ГБД (ЛДГ 1, 2) для перевода «тупикового» лактата в полифункциональный пируват.
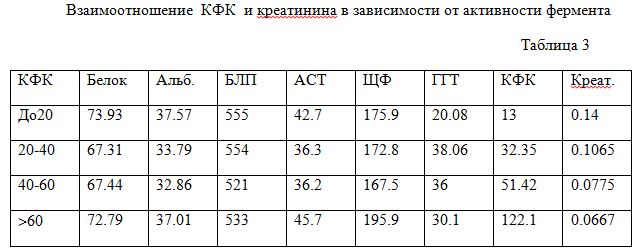
Хорошей иллюстрацией не простой корреляционной, а метаболической (точнее фермент-субстратной зависимости) является группировка показателей нескольких крыс по уровню активности КФК (табл.3). Повышение уровня КФК происходит на фоне снижения креатинина, появляющегося из дефицитного креатина. Выделенная зависимость отмечается при стабильном уровне альбумина, БЛП и АСТ (метаболический покой) и только ЩФ с ГГТ выявляют поздний и ранний подъем при повышении КФК.
Ферменты являются одним из колебательных контуров живых систем, создающим стабильность основного биохимического параметра крови – общего белка. Ферменты обеспечивают стабильность важнейших констант крови – общего белка, альбумина, глюкозы, холестерина, мочевины и креатинина. «Постоянство внутренней среды организма есть необходимое условие свободной жизни» (К.Бернар). Этот закон сохраняется на уровне биохимических показателей крови. Только с учетом уровня общего белка возможно отслеживать нацеленное (точечное) воздействие экзогенных факторов. Мощность ферментных систем должна обеспечивать обновление белков печени наполовину за 10 суток, у инертных коллагеновых белков период обновления составляет 300 дней. Крысы с более высоким уровнем активности ферментов имеют средний период полуобновления белков в пределах 3.5 суток. Коэффициент изнашивания (суточная потеря) по Рубнеру составляет 23 г белка на 70 кг веса. В сутки при этом выделяется 20-30 г мочевины при содержании в крови – 20-30 мг%. 100-120 г белка в сутки необходимо при общем калораже в 2500 ккал и норма белка повышается на 10 г при увеличении калоража на 500 ккал. Мощность ферментов переваривания на примере пепсина составляет 50 кг белка за 2 часа 1 г пепсина. Все ткани организма используют аминокислоты крови – 32-55 мг% - и накачивают их против градиента концентрации при участии ГГТ, т.к. в тканях их содержание значительно выше – до 500 мг%. Связано это не только c необходимостью использовать их для синтеза белка, но и стабилизировать их структуру в противовес мочевине. Однако только глицин, пролин и аланин не влияют на скорость ферментативных реакций и поэтому для них должна быть избирательность. Глицин создает микроклеточную буферную среду для улучшения растворимости разных веществ. В лабораториях трис-глициновый буфер является оптимальным буферным раствором для экспериментов. В сутки организм использует до 30 г аспарагиновой кислоты и до 40 г глицина, что не может быть обеспечено пищевыми белками. Физиологическая мощность альбумина – 1 г удерживает 18 г воды.
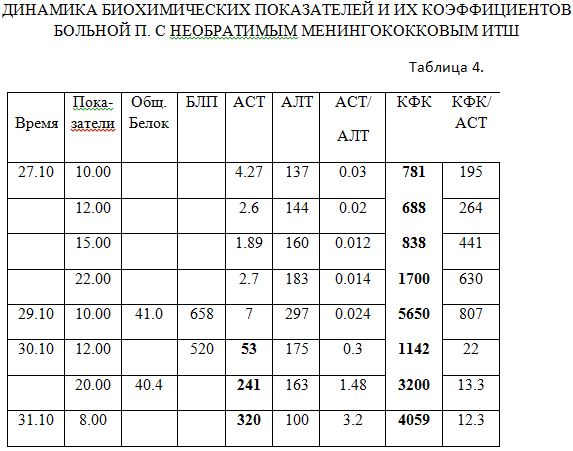
Представленная почасовая динамика уникальна (табл.4) удивительным несоответствием активности двух трансаминаз и КФК, а главное, их разновременностью, что подчеркивает вплоть до критического срока (29.10 в 10.00) их нецитолитический смысл. На фоне нулевых значений АСТ КФК продолжает повышаться, но «печеночная» АЛТ достигает максимума и это является началом угрожающего цитолиза по АСТ. Такие уникальные ферментативные сдвиги происходят на фоне резкой гипопротеинемии и токсикоза с повышением лизосомальной бета-глюкуронидазы. Наработка неотона происходит с первых анализов значительно и мощно. Повышение АЛТ – это глюкокортикоидный след адаптации особенно на фоне снижения белка, а более центральная зона метаболизма (АСТ это индикатор митохондриальной энергетики) изолирована за счет артерио-венозного шунтирования и практически не определяется в первых анализах. Всё просто и понятно с точки зрения патогенетических явлений, а не цитолитической простоты. И именно в клинике даже минимальным значениям КФК нужно уделять пристальное внимание как сигнал из зоны глубинной адаптации (а она в сущности всегда глубинна!)
