





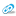
Отклонения от некого стандарта – это есть начало гомеокинеза, иначе восстановления гомеостаза (К.Бернар) с выраженной биохимической динамикой. И как это ни парадоксально звучит – основной результирующий признак гомеостаза – постоянная температура тела, которая складывается из двух составляющих: термогенеза и термопотребления (в виде собственно тепла и конкретно-химической ипостаси – АТФ), а патогенетические механизмы и являются биохимическими сдвигами.
На организменном, органном, тканевом, клеточном и субклеточном уровне антагонизм морфологических элементов является основой их взаимоотношений в режиме энергетической достаточности и субстратной наполненности. Эмбриональная стволовая клетка как «онтогенетический консервант» полностью развертывает свою программу в онтогенезе под влиянием крови в условиях субстратно-кислородного равновесия. Известные нам показатели могут (и несут) важнейшую метаболическую информацию стержневого типа для оценки энергетически глубинного статуса. Энергетическая стоимость физиологических функций рассматривается от кровообращения до обновления молекулярного состава организма, а свойства крови функционально емки и информативно понятны. Именно как интегрирующая среда кровь должна иметь наибольшую стабильность по всем параметрам. Отклонение минимального, а не статистически достоверного характера именно в контексте энергетического единства подчинено иерархии биохимических показателей крови.
В клинической биохимии великолепная семерка (АСТ, АЛТ, ЩФ, ГГТ, ЛДГ, ГБД и КФК) ферментов осталась неприкосновенной и в ней заключена суть, прежде всего суть метаболических качеств ферментов, подчиненных энергетической сущности живого, что и является одним из проявлений общего адаптационного синдрома. Конечная, промежуточная и начальная точка сдвигов подчинена естественному финалу – нормальному биологическому окислению (т.е. тканевому дыханию). И ежесекундная связь с внешним миром через кислородное потребление означает необходимость выстраивать небесно-земную (а точнее энерго-материальную) вертикаль и как это не парадоксально звучит, у организма на каждом уровне должны быть опознавательные признаки всей этой энергосущности. И возглас воинов Чингисхана: «Небо поможет!» лишь подчеркивает это неразрывное духовно-материальное (иначе субстратно-энергетическое) единство. И как не вспомнить при этом законченное гением обобщение: «Есть на земле предостаточно рас – просто цветная палитра, - воздух каждый вдыхает за раз два с половиной литра!» (В.С.Высоцкий). Однако в семерке ферментов у ЩФ, ЛДГ и ГБД – физиологическая (интервальная) манера поведения, АСТ, АЛТ, КФК обнаруживают свое поведение по закону «все или ничего!», а ГГТ имеет шаговое или интервальное присутствие в крови. И это не случайно! Однако количественная и качественная характеристика этих показателей в информативном отношении объясняет фундаментальные сдвиги всего метаболизма в целостном организме. Константность метаболических (белка, глюкозы, холестерина, креатинина) показателей обеспечивается вариативностью основных энзимологических параметров именно в цельной крови за счет субстратно-энзимологической оптимальности гармонического характера и биоритмологического качества. Мочевина несколько выпадает из этого ряда по причине катаболического исхода.
Статистика для ферментов непригодна, так как у них разное по выраженности автономное поведение подчинено поддержанию глюкозы на оптимальном уровне, резервированию адаптивных (глюкокортикоиды), половых (эстрогены, андрогены) и физиологических (минералокортикоиды) возможностей в виде общего энергоемкого предшественника (холестерина) на все случаи жизни и состояния. Для нас же в современном понимании – холестерин – это лишь субстрат тревоги за утраченное здоровье (ожирение, диабет, атеросклероз). Глюкоза, холестерин, общий белок являются физиологически интервальными и сходными с показателями дыхания, пульса и давления. Статистика старается «отстраниться» от любых колебаний в цифровом поле поиском доверительного интервала у получаемых показателей. Однако никаких колебаний быть не должно, а ожидание цифровой инертности означает сдерживание в ходе предварительных и окончательных выводов.
Статистические ворота в зоне энзимологической безразмерности это и есть цена адаптации не в филологическом, а конкретно-химическом смысле в каждом звене и каждой метаболической зоне. Тем более что стабильность глюкозы подчинена динамике метаболически единому именно по энергетической сущности холестерину, несмотря на его некоторую метаболическую отстраненность. Числовое равенство содержания глюкозы и холестерина (около 5 ммоль/л) также является порукой физиологического равноправия и абсолютной метаболической съединенности. И погружение в глубинные теоретические основы биохимии лишь подчеркивает энергетическое равенство и сходный энергетический потенциал: сгорание глюкозы в аэробном гликолизе – 38 молекул АТФ, а структура холестерина «обкрадывает» организм на (15 ацетилов-КоА х 12 АТФ =) 180 молекул АТФ. Мысленно, а значит и адекватно, мы выявляем доказательства правильности выбранного аналитического подхода.
Поэтому надпочечники, маленькие и анатомически незримые, хотя и содержат весь энергофонд организма в упакованном и директивном (роль гормонов не надо объяснять!) в своем стартовом смысле подчиняются математически точной интерпретации. Это позволяет выявлять специфические признаки гармонии любых организмов и состояний в режиме соотношения метаболических блоков, и поиск таких зависимостей очень ценен и продуктивен. Глюкоза является вершиной метаболических процессов и поддержание приоритетного биохимического (он же физиологический) параметра обеспечивается фондом белка, колебания которого и численно (4,5-5,5 для глюкозы и 75-85 для белка) и количественно (ммоль/л для глюкозы и г/л для белка) несопоставимы. Колебания ее подчас сопряжены именно с активацией метаболических путей для поддержания определенного уровня физиологического благополучия без формирования патологии углеводного обмена. Глюкоза необходима для настройки всего метаболизма в первую очередь!
Большой потенциал (фонд) белков обеспечивает узко специализированный и, в сущности, ограниченный фонд глюкозы. И ее результирующее воздействие в рецепторных зонах нервной и иммунной систем качественно стабильно, но количественно неосязаемо, т.к. интенсивность поступления ее в ткани не учитывается. Поэтому только ферменты могут указывать на интенсивность ее поглощения, использования и перераспределения. А ведь в этом и заключается метаболическая приоритетность белка, холестерина и глюкозы. У белков грубая как материнство земли материальность с выходом на срочный (глюкоза) и отсроченный (холестерин с его производными) и только в некотором смысле «способ существования» (Ф.Энгельс). Это первопричина и первооснова всего живого и вечного. Марксистская ограниченность наглядно вылезает и на сугубо биологической территории.
У мочевины отмечаются значительные отклонения с нулевыми и резко повышенными значениями в финале катаболизма как признак богатства и абсолютного истощения организма. У алкоголиков в реанимации нередко содержание мочевины равно нулю! Однако такая вариативность опять же является следствием частичной и минимальной доли азота именно в белках. И здесь мы также оперируем малыми концентрациями в ммоль/л. Каждая молекула азота в организме на счету, а его основной источник – белок – нами потребляется не ежедневно, недостаточно и неполноценно. А креатинин, родственник мочевины по почечной зависимости подобно Каину и Авелю связан с ним одной цепью, но не одной целью. Он константен, определяется в мкмолевых количествах, регулирует биоэнергетику на уровне митохондрий и требует особого внимания на минимальные отклонения как признак масштабных изменений подобно перемещению слоев земной коры, а не только как показание для почечного диализа.
Поддержание физиологического уровня глюкозы в условиях фактического «голода» требует перестройки метаболических процессов в организме в режиме интенсивного ГНГ. Оба этих процесса зависят от активности термогенной АСТ и глюконеогенной АЛТ на стадии усиленного ГНГ. Переаминирование в этом случае должно многократно интенсифицироваться за счет трансаминаз с использованием углеродных скелетов аминокислот. Глюкоза является приоритетным параметром и интенсивность ГНГ должна быть преобладающей, но только не в относительном и абсолютном покое. Подтверждением этого является гипопротеинемия и гипоальбуминемия. Соотношение трансаминаз определяет соотношение ката- и анаболизма, т.е. отражают физиологическую координированность, которая в норме равна 1,5, а ферментемия по АСТ - это один из первых важнейших индикаторов всей биоэнергетики организма. И тут мы находимся в зоне распределения биологического потенциала от генетических глубин (каждая пара оснований в ДНК требует 3 молекул аммиака) до энергетического комфорта (углеродные скелеты аминокислот эффективно поддерживают необходимый ТГ).
Метоболическое благополучие (белок от 75 г/л и выше) достигается энзимологическими сдвигами, и это хорошо иллюстрирует отношение трансаминаз к ГГТ, минимальное именно в критической (ниже 65 г/л) зоне общего белка. Вирусный гепатит как уникальная модель таких изменений в зоне предельно допустимого физиологического уровня указывает на способность организма поддерживать важнейшие физиологические константы за счет конкретных физиологических механизмов на удивительном, по сути, статистическом уровне. «Аварийный» уровень трансаминаз и их отношение при этом абсолютно константно – 0,58 – для всех уровней общего белка и является прямым доказательством физиологической координированности в зоне ката- и анаболизма и их соотношение (АСТ/АЛТ) является индикатором глубины нарушенного баланса. Соотношение АСТ+АЛТ/ГГТ – это следующий параметр обеспеченности аминокислотами для катаболизма в ЦТК или анаболизма в ГНГ с обязательным соответствием уровню общего белка – 2,05 – 2.49 – 2.64, т.е. уровню до 65 – от 65 до 75 – и больше 75 г/л. Соотношение трансаминаз и это признак физиологической и патогенетической координированности ката- и анаболизма, а их суммарное отношение к ГГТ - это использование тканевого резерва. Три привычных для лаборатории фермента меняют свою активность в угоду поддержания константного для любого живого существа уровня общего белка. единство траты (индикатор АСТ), ресинтеза (индикатор АЛТ) и перераспределения (индикатор ГГТ) ложатся на базу белкового благополучия при индикаторной роли константной глюкозы. Приоритетная фундаментальная метаболическая характеристика различных физиологических и клинических состояний, а также видовых признаков должна быть законом в аналитическом движении в системе правильных координат .
Трансаминазы вместе с ГГТ нормализуют метаболические показатели крови. При ВГ для ГГТ характерна разноуровненность значений в значительно более «узком» диапазоне (до 1200 МЕ/л), чем значения АЛТ и АСТ. Более низкие значения ее активности по сравнению с трансаминазами и недостаточны для обеспечения компенсаторного трансаминирования. При других инфекциях используются разные источники или пулы аминокислот и их диагностический поиск в лабораторных параметрах требует иных представлений. Обеспеченность аминокислотами для ТГ и ГНГ за счет тканевого пула должна оцениваться через соотношение АСТ+АЛТ/ГГТ, т.е. субстратов для катаболизма в ЦТК или анаболизма в ГНГ при жесткой их координации. И также как соотношение трансаминаз – это признак физиологической и патогенетической координированности ката- и анаболизма, так и их суммарное отношение к ГГТ указывает на эффективность вспомогательного резервного тканевого транспорта. Нам представляется возможным более ясно обосновать доказательную ценность выявленного, так как традиционное упаковывание первичного материала в статистическую комфортность является очень серьезной этому помехой.
И для доказательства описанных механизмов мы провели разделение показателей сыворотки больных в зависимости от уровня активности ГГТ. При этом оказалось (табл. № 1), что на фоне одинаковых значений белка повышение уровня активности ГГТ в выбранных интервалах противоположно снижению уровня трансаминаз. Самая низкая тимоловая проба (44,5) при максимальной (792 МЕ/л) активности ГГТ и интенсивности ТГ и ГНГ (уровень активности АСТ и АЛТ) при этом минимальная. Максимум активности ГГТ приходится при минимуме денатурированного фонда белков. Отношение трансаминаз скоординировано с интенсивностью тканевых потоков. Указанные уровни активности ферментов физиологически закономерны и патогенетически объяснимы по сравнению с нормой. Отношение трансаминаз снижается при повышении ГГТ, что указывает на превалирование ГНГ при интенсификации потока аминокислот. Таким образом, стабильность уровня общего белка обеспечивается координированным изменением уровня активности важнейших в физиологическом отношении метаболических потоков при жесткой корреляционной зависимости функционального характера.
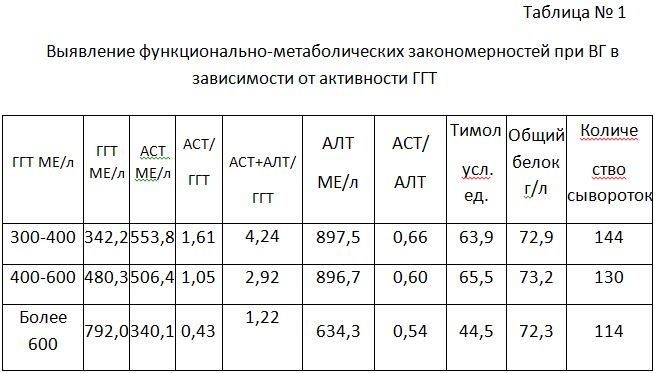
Распределение глюконеогенной АЛТ по валу автономно и количественно приоритетно, т.к. термогенез приоритетен в острой фазе ВГ, а источник субстратов (тканевые аминокислоты) при этом один. Эта реципрокность доминирующих процессов термогенеза (АСТ) и транспорта субстратов (ГГТ) координирована, что подтверждается их отношением. При этом регистрируется шаговая динамика – 1,61 – 1,05 – 0,43 – отношения двух ферментов как маркеров двух физиологических процессов. Очень сходна динамика коэффициента суммы двух трансаминаз по отношению к ГГТ: 4,24 – 2,92 – 1.22, что указывает на зависимость двух процессов от интенсивности поступления дефицитных аминокислот. Биохимический противоход – это и есть закон сохранения энергии и материи на единой субстратной основе.
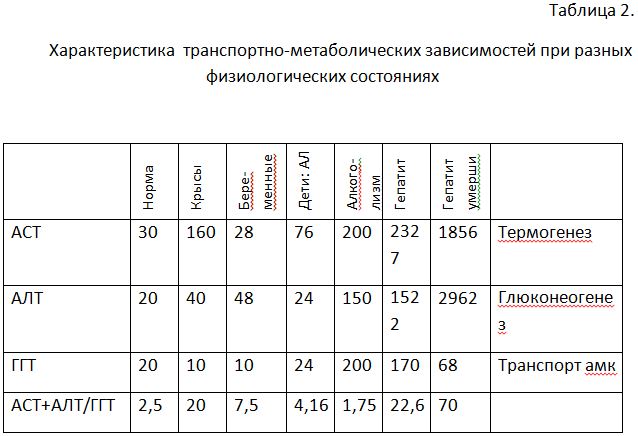
Физиологические особенности у разных объектов разные. Крысы: более свободный обмен катаболического типа, интенсивный ТГ, постоянное питание с элементами каннибализма, поставляющая аминокислоты ГГТ низкая. Беременные: физиологический дефицит белков, в последний триместр синтезируется до 1,5 тонн белка, ферментемия жесточайшей экономии атрофического типа с усилением ГНГ – анаболический (печеночный) тип отношения АСТ/АЛТ при минимальной ГГТ для сохранения соматической матрицы. Дети (астраханская лихорадка): ТГ превалирует над ГНГ – катаболический (сердечный) тип отношения АСТ/АЛТ с сохранением соматической белковой матрицы – ГГТ снижен. Алкоголизм: синдром «эндогенного выжигания белков» (катаболический уровень АСТ с компенсаторным уровнем АЛТ) при использовании последнего резервного (тканевого) пула – ГГТ - всегда повышена. Гепатиты: подпитка из тканей обязательна для выживания и соответствующее падение при терминальных состояниях. Снижение коэффициента указывает на увеличение доли тканевого фонда аминокислот без физиологической целесообразности, а увеличение - на дефицит белка для ГНГ и ЦТК. ГГТ транспортирует аминокислоты и пептиды в клетки, участвует в системе обезвреживания, а также в метаболизме биогенных аминов и все три механизма функционирования ГГТ существенны для любой патологии и особенно для гепатита. Количественный вклад каждого неодинаков, но на протяжении всего периода заболевания гепатитом утилизация денатурированных белков должна быть доминирующей.
ГГТ играет роль посредника и активного переносчика аминокислот между белковыми пулами и трансаминазами, как заказчиками интенсивности ката-, анаболизма. А работает она в системе оптимального дозирования по принципу «шаговых закономерностей». И переход на новый уровень интенсификации потока АМК детерминируется индивидуальным соотношением ката- и анаболизма.

Известно, что адаптационный синдром по Селье предполагает использование известных механизмов – ГНГ, протеолиза для поддержания важнейших биохимических констант, то есть для сохранения постоянства внутренней среды. Однако в противовес ему существует и закон отклонения гомеостаза как один из важнейших принципов регуляции метаболизма. Любая адаптивная реакция, как в норме, так и в патологии, сопровождается выбросом различных гормонов: адреналина, глюкокортикоидов и гормонов щитовидной железы, обеспечивающих существенные биохимические сдвиги, одним из проявлений которых является ферментемия.
А тройка ферментов – это символ равновесия в абсолютно едином масштабном представлении как обязательный признак вечно живого. И последнее: сроки жизни ферментов адаптации (АЛТ и ГГТ) более длительны, чем фермента базовой (АСТ) биоэнергетики.
